近日,重点实验室陈捷凯课题组研究发现RNA m6A修饰调控异染色质形成的新机制,阐明RNA m6A阅读器YTHDC1在这一机制中的关键作用:抑制基因组中广泛分布的ERVK、IAP、LINE1等转座元件,限制胚胎干细胞向全能性干细胞转化,相关研究论文以The RNA m6A reader YTHDC1 silences retrotransposons and guards ES cell identity为题于北京时间3月4日发表在Nature上。
RNA上的N6-腺苷酸甲基化(m6A)是mRNA上最丰富的RNA修饰,YTH结构域是已知的能够特异性识别m6A的结构域,因此RNA m6A可通过被含YTH结构域的阅读器蛋白(reader)识别进而参与各种生物学功能。通过基因敲除实验科学家发现,m6A催化酶METTL3敲除的小鼠在E8.5前就胚胎致死,而大部分m6A阅读器基因的敲除并不导致胚胎致死,包括三个分布在细胞质中主要参与mRNA调控的YTH基因(Ythdf1-3),只有分布在细胞核中的Ythdc1敲除后会出现早期的胚胎致死,提示m6A可能具有与YTHDC1有关的其他尚不清楚的重要生物学功能。
由于文献中至今尚未报道能够建立Ythdc1敲除的胚胎干细胞系,陈捷凯课题组运用条件敲除策略,针对编码YTH结构域重要部分的Ythdc1外显子7-9进行条件性敲除,发现敲除后细胞增殖能力迅速下降,基因表达谱出现2C-like(2C指受精卵卵裂后的二细胞期,小鼠在这一阶段启动合子基因组激活)的特征基因激活和逆转录转座元件激活。胚胎干细胞属于多能性干细胞,在小鼠胚胎发育中与囊胚内细胞团(ICM)的发育阶段相近,而2C-like细胞则具备全能性,多能性干细胞在培养中会随机出现少量的2C-like细胞,这一多能性-全能性的转变过程是近年来干细胞领域的研究热点,科学家希望通过了解其机制,发现维持全能性的可能方法。此前科学家曾发现RNA m6A调控胚胎干细胞退出多能性,这一工作进一步发现RNA m6A在全能性-多能性转变中的调控作用。
课题组在之前的工作中曾发现H3K9me3催化酶SETDB1在体细胞重编程、转座元件抑制、全能性-多能性转化中的重要作用(Nature Genetics 2013,Cell Reports 2020),因为表型一致,他们进一步研究RNA m6A是否参与了SETDB1-H3K9me3的调控。
遗传物质以染色质的形式存在,其中活跃转录的部分为常染色质,致密的异染色质代表被沉默的遗传信息。组蛋白H3K9me3是异染色质的重要修饰,也是细胞命运决定的重要机制。在小鼠胚胎干细胞(mES细胞)中,除了组成型异染色质(着丝粒、端粒等),H3K9me3主要标记遍布基因组的逆转录转座子(Retrotransposons)上并沉默这些区域。逆转录转座子上的H3K9me3由SETDB1负责催化,敲除SETDB1会导致这些元件重激活并使胚胎干细胞转化为具有全能性特征的、类似2细胞期的细胞(2C-like细胞)。但SETDB1是如何特异性地被募集到逆转录转座子区域的?目前有一些机制研究揭示,哺乳动物进化过程中会通过含KRAB结构域的锌指蛋白的进化来特异性沉默一部分转座元件(KRAB结构域会通过KAP1招募SETDB1),但这个机制并不覆盖所有各种类型的转座元件,提示还存在未发现的机制。
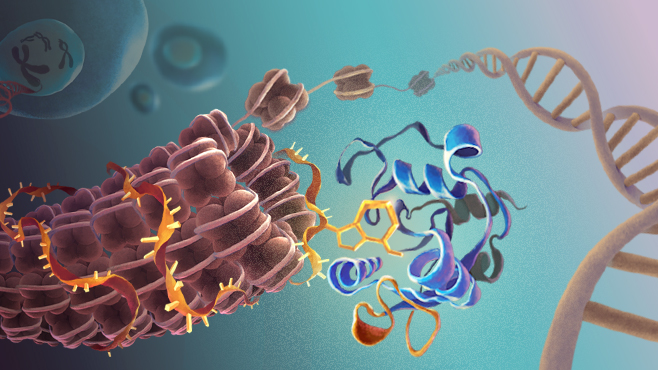
科研人员发现,YTHDC1能够直接结合转座子元件(TE)所转录出来的RNA上的m6A修饰,并招募SETDB1到相应的染色质位置,催化转座元件上的H3K9me3、形成异染色质并使这些转座元件沉默。敲除Ythdc1,会导致Setdb1依赖性的H3K9me3信号大幅度下降,证明YTHDC1是SETDB1介导逆转录转座子沉默的重要机制环节,也揭示RNA m6A调控染色质的功能。
RNAm6A结合蛋白YTHDC1调控异染色质形成